图7-31 ATP合酶的形态
(a) 电镜照片; (b)根据电镜照片绘制的模式图和各部分的大小。
● 组成
F1颗粒是一个多组分的结构,将它称为F0F1 ATP酶复合物, 或ATP合酶,在分离状态下具有ATP水解酶的活性,在结合状态下具有ATP合成酶的活性。除了线粒体中有ATP合酶外,植物叶绿体的类囊体和好氧细菌都有ATP合酶的同源物,线粒体ATP合酶有F0和F1两部分组成(图7-32), 主要功能是进行ATP的合成。也有学者将它看成是线粒体内膜呼吸链的第五个复合物(complex Ⅴ)。
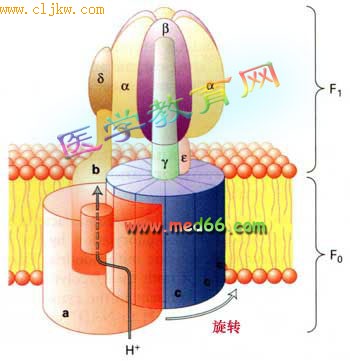
图7-32 ATP合酶的结构和组成
■ 氧化磷酸化偶联机理:化学渗透假说
关于氧化磷酸化的偶联机理,先后提出过几种假说,如化学偶联假说(chemical coupling hypothesis)和构象偶联假说(conformational coupling hypothesis),这些假说由于证据不足得不到公认。
英国生物化学家P.Mitchell 于1961年提出了化学渗透偶联假说(chemiosmotic coupling hypothesis)解释氧化磷酸化的偶联机理。该学说认为:在电子传递过程中, 伴随着质子从线粒体内膜的里层向外层转移, 形成跨膜的氢离子梯度,这种势能驱动了氧化磷酸化反应(提供了动力), 合成了ATP.这一学说具有大量的实验支持,得到公认并获得了1978年诺贝尔奖。化学渗透学说可以很好地说明线粒体内膜中电子传递、质子电化学梯度建立、ADP磷酸化的关系(图7-33)。
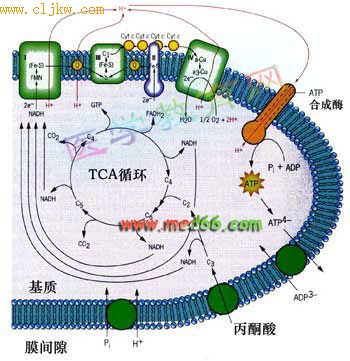
图7-33 线粒体在有氧呼吸中的主要作用
■ ATP合酶合成ATP的机理
● 结合变构模型(binding-change model)
ATP合酶合成ATP的分子机理的研究一直是研究的热点, 为多数人接受的ATP合酶合成ATP的模型是“结合变构模型”。该模型认为F1中的γ亚基作为C亚基旋转中心中固定的转动杆, 旋转时会引起αβ复合物构型的改变。有三种不同的构型,对ATP和ADP具有不同的结合能力: ①O型几乎不与ATP、ADP和Pi结合;②L型同ADP和Pi的结合较强;③T型与ADP和Pi的结合很紧,并能自动形成ATP,并能与ATP牢牢结合。当γ亚基旋转并将αβ复合物转变成O型则会释放ATP(图7-34)。
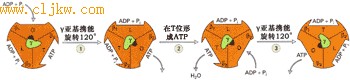
图7-34 结合变构模型
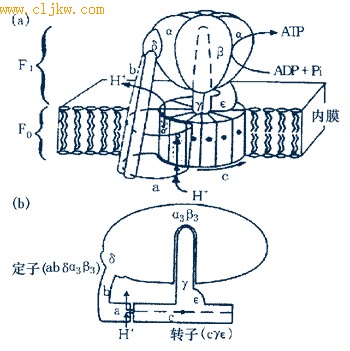
● 定子(stator)和转子(rotor)
Timothy等人提出了一个ATP合酶中能量转化过程的模型(图7-35),该模型认为由abα3β3δ组成了“定子(stator)”,cγε则形成“转子(rotor)”。当H+质子穿过a和c之间的通道时产生了力矩,从而推动了转子与定子间的相对转动,这样在F1中合成了ATP.
上一页 [1] [2] [3] 下一页
