溶酶体的生物发生
溶酶体的形成是一个相当复杂的过程, 涉及的细胞器有内质网、高尔基体和内体等。比较清楚的是甘露糖-6-磷酸途径(mannose 6-phosphate sorting pathway):溶酶体的酶类在内质网上起始合成, 跨膜进入内质网的腔, 在顺面高尔基体带上甘露糖6-磷酸标记后在高尔基体反面网络形成溶酶体分泌小泡, 最后还要通过脱磷酸才成为成熟的溶酶体(图9-44)。
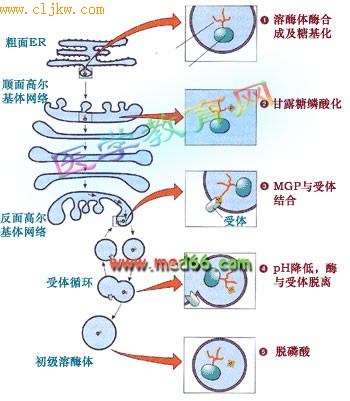
图9-44 溶酶体的酶寻靶过程、涉及的细胞器及机理
大多数溶酶体的酶在寡糖链上含有甘露糖, 在顺面高尔基网络转变成甘露糖-6-磷酸。新形成的溶酶体的酶通过高尔基复合体,在高尔基体反面网络与膜受体结合后被包进溶酶体分泌小泡,通过出芽形成自由的分泌泡。通过H+-质子泵调节溶酶体分泌小泡中的pH,使溶酶体的酶同受体脱离,受体再循环, 溶酶体酶脱磷酸后成为成熟的初级溶酶体。
■ 溶酶体酶蛋白的M6P标记
研究发现,溶酶体的酶上都有一个特殊的标记∶6-磷酸甘露糖(mannose 6-phosphate, M6P)。这一标记是溶酶体酶合成后在粗面内质网和高尔基体通过糖基化和磷酸化添加上去的。
● 糖基化
溶酶体酶蛋白在膜旁核糖体上合成,进入内质网后进行N-连接糖基化, 经加工后形成带有8个甘露糖残基和2个N-乙酰葡萄糖胺残基的糖蛋白转运到高尔基体。
● 信号斑(signal patch)
信号斑是溶酶体酶蛋白多肽形成的一个特殊的三维结构, 它是由三段信号序列构成的, 可被磷酸转移酶特异性识别(图9-45)。
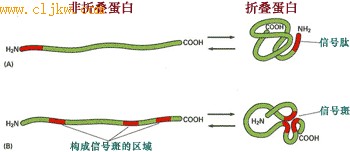
图 9-45 信号斑
溶酶体蛋白的多肽上有三段信号序列, 通过折叠,三个信号序列相互靠近形成信号斑结构。
● 甘露糖磷酸化的酶
将磷酸基团添加到溶酶体酶的甘露糖的第六位碳上的反应是由两种酶催化的,一种酶是N-乙酰葡萄糖胺磷酸转移酶(N-acetyglucosamine phosphotransferase),另一个酶是N-乙酰葡萄糖苷酶, 功能是释放N-乙酰葡萄糖胺(46)。
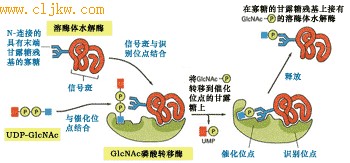
图9-46 溶酶体酶蛋白信号斑与磷酸化酶相互作用
顺面高尔基体中的N-乙酰葡萄糖胺磷酸转移酶有两个功能位点,一个识别位点能够识别信号斑并与之结合;催化位点与高甘露糖N-连接的寡聚糖以及UDP-GlcNAc结合。
● 磷酸化反应
反应中磷酸基的供体是UDP N-乙酰葡萄糖胺(N-acetyglucosamine,GlcNAc), 甘露糖残基磷酸化的位点是第六位碳原子(图9-47)。每个溶酶体酶蛋白至少有一个甘露糖残基被磷酸化。
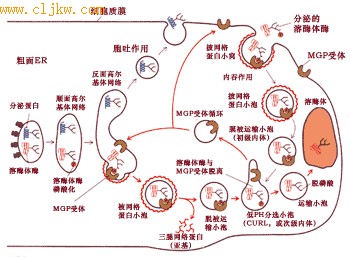
图9-47 溶酶体酶蛋白甘露糖残基磷酸化的生化反应
■ 溶酶体酶的M6P分选途径
这一途径的两个关键是:M6P标记和M6P受体蛋白。
● M6P受体蛋白(M6P receptor p
[1] [2] [3] 下一页
